Сложное строение человеческого мозга открывает тайны нашего поведения, объясняет законы мыслительной деятельности, протекание эмоций и чувств. Каждое полушарие головного мозга отвечает за свои специфические функции и задачи (например, известно, что правое отвечает за логику, а левое - за воображение и фантазию), но имеются также структуры, которые обеспечивают единую и согласованную работу всей центральной нервной системы. Одной из таких структур является ретикулярная формация.
Общие сведения
Ретикулярная формация - это участок стволового отдела мозга, представленный разветвленной сетью нервных клеток и ядер, соединяющих между собой различные отделы головного мозга. В отличие от других структур, - например, таламуса, гипоталамуса, мозжечка, - которые имеют некую цельную форму (ядра, железы), ретикулярная формация не представлена единым морфологическим образованием, а является «сетью» (от латинского reticulum — сеть) дендритов и аксонов, которые с разной степенью плотности проникают между отделами и структурами головного мозга, объединяя их между собой и обеспечивая их совместную деятельность.
Метафорически выражаясь: если наш мозг представить в виде некоего изделия, скажем, рубашки, то ретикулярная формация - это нити, которыми рубашка сшита. Ретикулярная формация пронизывает структуры продолговатого, среднего мозга и моста, имеет непосредственные связи с мозжечком, спинным мозгом, таламусом и опосредованные - с вышележащими отделами: гипоталамусом, зрительными ядрами и корой.
Как устроена
В состав ретикулярной формации входит огромное количество нейронов с разветвленными дендритами и длинными аксонами, за счет чего становится возможным передавать нервные импульсы в различные отделы головного и спинного мозга. При этом можно выделить две наиболее крупные группы нейронных скоплений:
- Ретикулотегментальное ядро, нейроны которого получают сигналы от вышележащих отделов ГМ (четверохолмие, таламус) и передают их далее в структуры мозжечка, регулируя тем самым некоторые жизненно важные двигательные функции: координация взгляда, движения глаз.
- Латеральное ядро, нейроны которого восходят от структур спинного мозга и вестибулярных ядер и обеспечивают информирование коры ГМ о положении тела в пространстве, участвуют в регуляции дыхания и сосудистой иннервации.
- Кроме того, в состав ретикулярной формации входят нейроны, которые принимают важное участие в работе центров терморегуляции, насыщения и голода.

Основные функции
Основное предназначение ретикулярной формации состоит в сенсорном анализе многочисленных сигналов, поступающих от различных отделов ГМ.
За счет тесных связей со спинным мозгом она также принимает самое активное участие в моторной регуляции, начиная от глотательного рефлекса и заканчивая сложными двигательными операциями. Кроме того, ретикулярная формация осуществляет активирующее воздействие на весь ГМ в целом, участвуя в регуляции циклов сна и бодрствования.
В общем виде функции ретикулярной формации сводятся к следующим:
- Регуляция скелетной мускулатуры (участвует в управлении движениями тела) и вегетативных функций (дыхание, чихание, кровообращение и т.п.).
- Контроль процессов сна и бодрствования (за счет оказания активирующего и тормозящего воздействия на кору больших полушарий).
- Активизирующая функция (проявляется в том, что ретикулярная формация обеспечивает постоянное тоническое возбуждение коры ГМ, за счет чего становится возможным поддержание внимания, сознания и протекание мыслительных процессов.)
- Обработка сигналов, поступающих из внешней и внутренней среды.
Отличительные особенности работы ретикулярной формации связаны, прежде всего, с определенными свойствами ее :
Нервные клетки, входящие в состав ретикулярной формации, обладают повышенной способностью к тоническому возбуждению. Это означает, что бо́льшая часть нейронов находятся в постоянном возбуждении и генерирует нервные импульсы, которые передаются в вышележащие отделы ГМ. Такая тоническая активность обусловлена рядом факторов:
- Проходимость большого количества сигналов через структуры ретикулярной формации. Приведем простую аналогию: представьте себе рояль или какой-либо другой струнный инструмент. Понятно, что когда мы напрямую касаемся струн, они начинают колебаться и издавать звук. То же самое происходит и с нервными клетками, когда к ним приходят сигналы от других нейронов.Однако представим далее, что мы не касаемся непосредственно струн инструмента, но, скажем, скачем рядом с ним, сильно стуча ногами о пол. Звука инструмента мы, может быть, и не услышим, но едва заметное колебание струн все же будет иметь место. То же самое происходит и с нейронами ретикулярной формации. Поскольку через нее постоянно проходят какие-то сигналы (как афферентные, так и эфферентные) от различных структур ЦНС, то это и создает постоянное тоническое возбуждение нейронов ретикулярной формации, за счет того, что она находится в эпицентре постоянного нервно-импульсного обмена.
- Повышенная чувствительность нейронов к химическим веществам (гормоны, медикаменты, психотропные вещества). Чашка кофе, выпитая утром, «включает» именно структуры ретикулярной формации и за счет длительного сохранения возбуждения в ее нейронах поддерживает нас в активном состоянии.
Нисходящее и восходящее влияние РФ
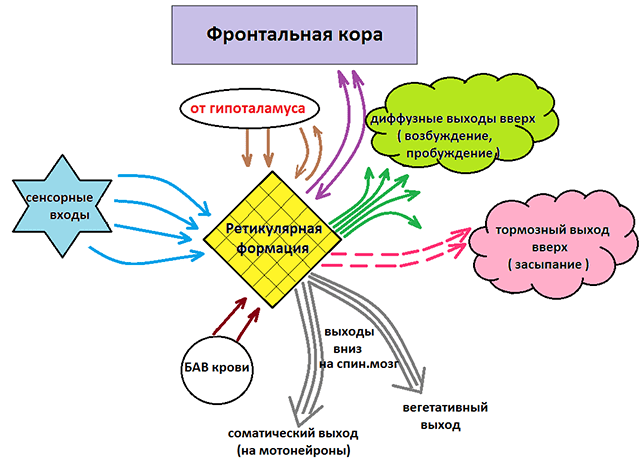
Как уже отмечалось, ретикулярная формация оказывает возбуждающее и тормозящее действие на различные отделы ГМ. При этом можно выделить два отдела, которые специализируются на передаче возбуждения в те или иные структуры мозга.
Нисходящий отдел: представлен вегетативными и двигательными центрами и оказывает нисходящее влияние на отделы спинного мозга. Соответствующие нейронные скопления регулируют деятельность дыхательного, сосудодвигательного, слюноотделительного центров, а также центров, отвечающих за построение простых и сложных двигательных реакций. Это указывает на решающую роль именно центральной нервной системы в регуляции даже элементарных безусловных рефлексов. Стимуляция нисходящего отдела приводит к затормаживанию спинальных центров и вызывает в естественной среде состояние глубоко сна (спать «без задних ног»). Тот же эффект можно вызвать и искусственным путем, например, вводя человека в состояние транса или наркоза.
Восходящий отдел: представлен нервными волокнами, соединяющими структуры ретикулярной формации с вышележащими отделами: таламусом, гипоталамусом, мозжечком и корой. Восходящее влияние оказывает стимулирующее действие на кортикальные структуры и обеспечивает активное состояние сознания. Восходящее влияние не прекращается, даже когда мы спим. Если бы наш мозг мог полностью «отключаться», то каждое пробуждение было бы сродни рождению: кто я? Где я? Как я сюда попал? Однако за счет работы ретикулярных структур у нас сохраняется возможность всегда возвращаться в то исходное состояние сознания, в котором мы находились до момента сна. Кроме того, и во время ночного отдыха у нас остается способность реагировать на некоторые жизненно важные раздражители, т.е. мы, как правило, не спим «мертвым сном» и можем проснуться, если рядом зашевелился и заплакал ребенок, что-то громко упало и т.п.
Проявление повреждения структур
Ретикулярная формация играет значимую роль в интегративной деятельности всего головного мозга. За счет выполнения функции ведущего проводника всех видов нервных импульсов во все отделы ЦНС, ретикулярная формация находится в постоянной работе. Чрезмерные умственные и эмоциональные перегрузки вредны для мозга в целом и для ретикулярной формации, в частности. К счастью, своевременный прием седативных препаратов может (за счет повышенной восприимчивости нейронов к химическому воздействию) быстро исправить ситуацию и нормализовать состояние.
Однако возможны и менее благоприятные исходы. Повреждение возможно в результате черепно-мозговых травм, онкологических заболеваний головного мозга, инфекционных поражений.
Основным проявлением неблагополучия является потеря сознания.
Нарушение восходящих связей обнаруживает себя в состоянии апатии, слабости, повышенной сонливости, двигательной растороможенности, нарушениях ночного сна. Нередки сопутствующие вегетативные расстройства.
Термин ретикулярная формация предложил в 1865 году немецкий ученый О. Дейтерс. Под этим термином Дейтерс понимал разбросанные в стволе головного мозга клетки, окруженные множеством волокон, идущих в различных направлениях. Именно сетевидное расположение волокон, связывающих между собой нервные клетки, послужило основой для предложенного названия.
В настоящее время морфологами и физиологами накоплен богатый материал о строении и функциях ретикулярной формации. Установлено, что структурные элементы ретикулярной формации локализуются в целом ряде мозговых образований, начиная с промежуточной зоны шейных сегментов спинного мозга (VII пластина), и заканчивая некоторыми структурами промежуточного мозга (интраламинарными ядрами, таламическим ретикулярным ядром). Ретикулярная формация состоит из значительного числа нервных клеток (в ней содержится почти 9/10 клеток всего ствола мозга). Общие черты строения ретикулярных структур - наличие особых ретикулярных нейронов и отличительный характер связей.
Рис. 1. Нейрон ретикулярной формации. Сагиттальный разрез ствола мозга крысенка.
На рисунке А представлен только один нейрон ретикулярной формации. Видно, что аксон разделяется на каудальный и ростральный сегменты, большой протяженности, со множеством коллатералей. Б. Коллатерали. Сагиттальный разрез нижней части ствола мозга крысенка, показывающий соединения коллатералей большого нисходящего пути (пирамидный путь) с ретикулярными нейронами. Коллатерали восходящих путей (сенсорные пути), отсутствующие на рисунке, соединяются с ретикулярными нейронами подобным же образом (по Шейбэлу М. Э. и Шейбэлу А. Б.)
Наряду с многочисленными отдельно лежащими нейронами, различными но форме и величине, в ретикулярной формации головного мозга имеются ядра. Рассеянные нейроны ретикулярной формации прежде всего играют важную роль в обеспечении сегментарных рефлексов, замыкающихся на уровне ствола головного мозга. Они выступают в качестве вставочных нейронов при осуществлении таких рефлекторных актов, как моргание, роговичный рефлекс и т. д.
Выяснено значение многих ядер ретикулярной формации. Так, ядра, расположенные в продолговатом мозге, имеют связи с вегетативными ядрами блуждающего и языкоглоточного нервов, симпатическими ядрами спинного мозга, они участвуют в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и т. д.
Установлена роль голубого пятна и ядер шва в регуляции сна и бодрствования. Голубое пятно , находится в верхнелатеральной части ромбовидной ямки. Нейроны этого ядра продуцируют биологически активное вещество - норадреналин , который оказывает активирующее воздействие на нейроны вышележащих отделов мозга. Особенно высока активность нейронов голубого пятна во время бодрствования, во время глубокого сна она угасает почти полностью. Ядра шва располагаются по срединной линии продолговатого мозга. Нейроны этих ядер вырабатывают серотонин , который вызывает процессы разлитого торможения и состояние сна.
Ядра Кахаля и Даркшевича , относящиеся к ретикулярной формации среднего мозга, имеют связи, с ядрами III, IV, VI, VIII и XI пар черепных нервов. Они координируют работу этих нервных центров, что очень важно для обеспечения сочетанного поворота головы и глаз. Ретикулярная формация ствола головного мозга имеет важное значение в поддержании тонуса скелетной мускулатуры, посылая тонические импульсы на мотонейроны двигательных ядер черепных нервов и двигательных ядер передних рогов спинного мозга. В процессе эволюции из ретикулярной формации выделились такие самостоятельные образования, как красное ядро, черное вещество.
По структурно-функциональным критериям ретикулярная формация делится на 3 зоны:
1. Медианную, расположенную по средней линии;
2. Медиальную, занимающую медиальные отделы ствола;
3. Латеральную, нейроны которой лежат вблизи сенсорных образований.
Медианная зона представлена элементами шва, состоящие из ядер, нейроны которых синтезируют медиатор – серотонин. Система ядер шва принимает участие в организации агрессивного и полового поведения, в регуляции сна.
Медиальная (осевая) зона состоит из мелких нейронов, которые не ветвятся. В зоне располагается большое количество ядер. Встречаются также крупные мультиполярные нейроны с большим числом густо ветвящихся дендритов. Они образуют восходящие нервные волокна в кору больших полушарий и нисходящие нервные волокна в спинной мозг. Восходящие пути связи медиальной зоны оказывают активирующее влияние (прямо или опосредованно через таламус) на новую кору. Нисходящие пути оказывают тормозящее влияние.
Латеральная зона – к ней относятся ретикулярные образования, расположенные в стволе мозга вблизи сенсорных систем, а также ретикулярные нейроны, лежащие внутри сенсорных образований. Основным компонентом этой зоны является группы ядер, которые примыкают к ядру тройничного нерва. Все ядра латеральной зоны (за исключением ретикулярного латерального ядра продолговатого мозга) состоят из нейронов малой и средней величины и лишены крупных элементов. В этой зоне располагаются восходящие и нисходящие пути, обеспечивающие связь сенсорных образований с медиальной зоной ретикулярной формации и моторными ядрами ствола. Эта часть ретикулярной формации является более молодым и возможно прогрессивнее, с ее развитием связан факт уменьшения объема осевой ретикулярной формации в ходе эволюционного развития. Таким образом, латеральная зона – это совокупность элементарных интегративных единиц, сформированных вблизи и внутри специфических сенсорных систем.
Рис. 2. Ядра ретикулярной формации (РФ) (по: Niuwenhuys еt. аl, 1978).
1-6 - медианная зона РФ: 1-4- ядра шва (1 - бледное, 2 - темное, 3 - большое, 4- мостовое), 5 - верхнее центральное, 6 - дорсальное ядро шва, 7-13 - медиальная зона РФ: 7 - ретикулярное парамедианное, 8 - гигантоклеточное, 9 - ретикулярное ядро покрышки моста, 10, 11 - каудальное (10) и оральное (11) ядра моста, 12 - дорсальное покрышечное ядро (Гуддена), 13 - клиновидное ядро, 14- I5-латеральная зона РФ: 14 - центральное ретикулярное ядро продолговатого мозга, 15 - латеральное ретикулярное ядро, 16, 17 - медиальное (16) и латеральное (17) парабрахиальные ядра, 18, 19 - компактная (18) и рассеянная (19) части педункуло-понтийного ядра.
Благодаря нисходящим влияниям ретикулярная формация оказывает тоническое влияние и на мотонейроны спинного мозга, что в свою очередь повышает тонус скелетной мускулатуры, совершенствует систему обратной афферентной связи. В результате любой двигательный акт совершается значительно эффективнее, осуществляет более точный контроль за движением, но чрезмерное возбуждение клеток ретикулярной формации может привести к дрожанию мышц.
В ядрах ретикулярной формации находятся центры сна и бодрствования, и стимуляция тех или иных центров приводит или к наступлению сна, или к пробуждению. На этом основано применение снотворных. В ретикулярной формации расположены нейроны, реагирующие на болевые раздражения, идущие от мышц или внутренних органов. В ней также расположены специальные нейроны, которые обеспечивают быструю реакцию на внезапные, неопределенные сигналы.
Ретикулярная формация тесно связана с корой больших полушарий, благодаря этому формируется функциональная связь между внешними отделами ЦНС и стволом головного мозга. Ретикулярная формация играет важную роль как в интеграции сенсорной информации, так и в контроле над деятельностью всех эффекторных нейронов (моторных и вегетативных). Она имеет также первостепенное значение для активации коры больших полушарий, для поддержания сознания.
Необходимо отметить, что кора полушарий большого мозга, и в свою очередь, посылает по корково-ретикулярным путям импульсы в ретикулярную формацию. Эти импульсы возникают в основном в коре лобной доли и проходят в составе пирамидных путей. Корково-ретикулярные связи оказывают либо тормозное, либо возбуждающее действие на ретикулярную формацию ствола головного мозга, они осуществляют корректировку прохождения импульсов по эфферентным путям (отбор эфферентной информации).
Таким образом, между ретикулярной формацией и корой полушарий большого мозга имеется двусторонняя связь, которая обеспечивает саморегуляцию в деятельности нервной системы. От функционального состояния ретикулярной формации зависит тонус мускулатуры, работа внутренних органов, настроение, концентрация внимания, память и т. д. В целом ретикулярная формация создает и поддерживает условия для осуществления сложной рефлекторной деятельности с участием коры полушарий большого мозга.
Ретикулярная формация начинается в срединной части верхних шейных сегментов спинного мозга и продолжается в центральных отделах продолговатого мозга, моста, среднего и промежуточного мозга. Она представляет собой скопления нейронов (ядра) с многочисленными сильно ветвящимися отростками, идущими в разных направлениях и образующими густую сеть. Между нейронами ретикулярной формации образуется очень много синапсов. От всех поступающих в таламус, а затем в кору больших полушарий афферентных путей отходят многочисленные коллатерали к ретикулярной формации, чем и обеспечивается ее восходящая активирующая деятельность. Ретикулярная формация также получает импульсы из мозжечка, подкорковых ядер, лимбической системы, которые обеспечивают эмоционально-адаптивные поведенческие реакции, мотивационные формы поведения.
В ретикулярной формации человека выделяют 48 ядер. Наиболее крупным является гигантоклеточное ядро, которое содержит гигантские нейроны, отсутствующие в других ядрах. Важным свойством нейронов ретикулярной формации является их высокая химическая чувствительность к различным гуморальным факторам и фармакологическим веществам, особенно к анестезирующим препаратам и так называемым успокаивающим средствам.
В ретикулярной формации продолговатого мозга располагаются жизненно важные центры регуляции дыхания, сердечно-сосудистой системы, деятельности пищеварительного тракта, центры рефлекторных актов, связанных с вестибулярными и слуховыми нервами.
Установлено, что ретикулярная формация по восходящим нервным путям оказывает возбуждающее влияние на кору больших полушарий , а по нисходящим путям – возбуждающее или тормозящее действие на деятельность спинного мозга (рис. 84). Ретикулоспинальные влияния играют важную роль в координации простых и сложных движений, в реализации влияний психической сферы на осуществление сложной двигательной поведенческой деятельности человека.
Было обнаружено, что электрическое раздражение гигантоклеточного ядра ретикулярной формации вызывает неспецифическое торможение сгибательных и разгибательных рефлексов, осуществляемых мотонейронами спинного мозга. Влияние ретикулярной формации на мышечный тонус передается по двум ретикуло-спинальным путям: быстропроводящему и медленнопроводящему. Импульсы, поступающие по этим путям, повышают активность гамма-мотонейронов спинного мозга, что в свою очередь возбуждает альфа-мотонейроны, и мышечный тонус увеличивается. Ретикулярная формация может выступать не только в роли регулятора возбудимости мотонейронов спинного мозга, но и принимать участие в процессах, связанных с поддержанием позы и организацией целенаправленных движений.
Активность самой ретикулярной формации поддерживается непрерывным поступлением импульсов, идущих от рецепторов тела. Важная роль в поддержании ее активности принадлежит гуморальным факторам, по отношению к которым она обладает высокой чувствительностью.
Благодаря работам Х. Мегуна и Дж. Моруцци были открыты восходящие, активирующие влияния ретикулярной формации на кору больших полушарий (рис. 84, А). Обнаружено, что ретикулярная формация участвует в регуляции сна и бодрствования. Раздражение и возбуждение ее с помощью вживленных в мозг электродов вызывает пробуждение у спящих животных. Эта поведенческая реакция пробуждения сопровождается учащением ритма электроэнцефалограммы в обширных областях коры больших полушарий. У бодрствующего животного подобное раздражение повышало уровень корковой активности, усиливало внимание к внешним сигналам и улучшало их восприятие. Разрушение восходящих путей от ретикулярной формации приводит к глубокому сну у бодрствовавших животных и уменьшению частоты колебаний электроэнцефалограммы.
Ретикулярная формация может оказывать и тормозное влияние на кору больших полушарий. Оно имеет место в случае длительной и монотонной работы. Например, в производственных условиях при работе на конвейере или в спорте при прохождении длинных и сверхдлинных дистанций.
В электрических проявлениях деятельности мозга активирующие влияния ретикулярной формации проявляются в виде возникновения частой асинхронной активности (десинхронизация), а тормозящие влияния – в виде медленных ритмичных колебаний (синхронизация).
Большинство нейронов ретикулярной формации являются полисенсорными, т.е. отвечают на различные раздражения: световые, звуковые, тактильные и т.д. Эти нейроны имеют обширные рецептивные поля, большой латентный период и слабую воспроизводимость реакции, что сильно отличает их от нейронов специфических ядер. В связи с этим нейроны ретикулярной формации относят к неспецифическим. Точно также восходящие пути ретикулярной формации называют неспецифическими, т.к. они направлены к обширным областям коры больших полушарий в отличие от специфических проекционных путей от органов чувств, идущих в конкретные зоны коры.
Ретикулярной формации принадлежит важная роль в механизмах формирования условнорефлекторных реакций организма. Она повышает активность вегетативных нервных центров , функционируя совместно с симпатическим отделом вегетативной нервной системы. Введение адреналина повышает тонус ретикулярной формации, в результате чего усиливается ее активирующее влияние на кору больших полушарий. Адреналин, выделяемый мозговым веществом надпочечников при эмоциях, действуя на ретикулярную формацию, увеличивает и удлиняет эффекты возбуждения симпатической нервной системы.
Благодаря наличию кольцевых связей в ретикулярной формации происходит взаимодействие афферентных и эфферентных импульсов , возможна их продолжительная циркуляция по кругу. Вследствие этого поддерживается определённый уровень возбуждения самой ретикулярной формации, а она в свою очередь поддерживает тонус и готовность к деятельности различных отделов центральной нервной системы. Активность ретикулярной формации находится под регулирующим влиянием коры больших полушарий (рис. 84, Б).
5.16. Функции мозжечка
Мозжечок расположен позади и чуть выше продолговатого мозга и варолиева моста под большими полушариями. Это надсегментарная структура, появляющаяся на ранних этапах филогенеза хордовых животных. Степень развития мозжечка определяется сложностью среды обитания и передвижения организма. Наибольшего развития мозжечок достигает у человека в связи с прямохождением и усложнением движений при трудовой деятельности. В то же время мозжечок не является жизненно необходимым органом. У людей с врожденным отсутствием мозжечка не наблюдается каких-либо серьезных нарушений движений, препятствующих их жизнедеятельности.
Мозжечок человека состоит из непарной средней части – червя и расположенных по обе стороны от червя двух полушарий. Поверхность мозжечка покрыта серым веществом, толщиной 1–2,5 мм, образующим его кору. Под корой находится белое вещество, в котором группами располагается серое вещество, представляющее собой скопления тел нейронов – ядра мозжечка.
Мозжечок выполняет проводниковую, рефлекторную и интегративную функции. По афферентным спинно-мозжечковым путям в мозжечок поступают импульсы от рецепторов кожи, мышц и сухожилий. От вестибулярных ядер продолговатого мозга по вестибуло-мозжечковым путям в мозжечок поступает информация о положении тела. Кора больших полушарий также посылает афферентные пути в мозжечок, среди которых наиболее важными являются кортико-мосто-мозжечковый и кортико-ретикуло-мозжечковый пути.
Эфферентные пути от мозжечка идут к спинному и продолговатому мозгу, к ретикулярной формации, красным ядрам среднего мозга, к промежуточному мозгу, коре больших полушарий и к подкорковым ядрам.
Мозжечок оказывает рефлекторные влияния на различные двигательные и вегетативные функции. Главное его значение заключается в дополнении и коррекции деятельности остальных двигательных центров. Мозжечок участвует: 1) в регуляции позы и мышечного тонуса; 2) в коррекции медленных целенаправленных движений и координации их с рефлексами поддержания позы; 3) в координации быстрых целенаправленных движений, осуществляемых по команде из коры больших полушарий.
Зона коры червя мозжечка главным образом связана с регуляцией позы, равновесия и мышечного тонуса. Промежуточная околочервячная зона коры мозжечка принимает участие в координации медленных целенаправленных движений с рефлексами поддержания позы.
Боковые участки коры, расположенные на полушариях мозжечка, участвуют в осуществлении быстрых целенаправленных движений. К полушариям мозжечка от ассоциативных областей коры больших полушарий поступает информация о замысле движения по афферентному кортико-мосто-мозжечковому пути, насчитывающему около 20 млн. нервных волокон. В полушариях мозжечка и зубчатом ядре мозжечка информация о замысле движения преобразуется в программу движения, которая в таламусе промежуточного мозга объединяется с программой, поступающей от подкорковых ядер, и далее посылается в двигательные зоны коры больших полушарий. От коры больших полушарий по нисходящим путям передаются команды к мотонейронам спинного мозга и происходит осуществление движений.
Мозжечок вносит в выполнение движений необходимые поправки, обеспечивая точность, ловкость и координированность движений. При нарушении функций мозжечка возникают различные двигательные расстройства: атония, астения, астазия, атаксия, асинергия, дисметрия, адиадохокинез, дезэквилибрия (рис. 85).
Атония характеризуется резким ослаблением мышечного тонуса. Она обычно сопровождается астенией – слабостью и повышенной утомляемостью мышц. Астазия проявляется в утрате способности мышц к длительному тетаническому сокращению, вследствие чего конечности и голова непрерывно дрожат и качаются, т.е. наблюдается тремор. Атаксия характеризуется нарушением координации движений, походки и др. При атаксии ноги широко расставлены, движения избыточны, из-за которых больного как пьяного бросает из стороны в сторону.
Асинергия проявляется в нарушении взаимодействия между двигательными центрами различных мышц. При этом происходит как бы распад программы движения и целостное движение состоит не из одновременных содружественных актов, а распадается на ряд простых движений выполняемых последовательно. Асинергия сочетается с дисметрией , или утратой соразмерности движения. При дисметрии наблюдается несоответствие между интенсивностью мышечного сокращения и задачей выполняемого движения, движения становятся размашистыми и неупорядоченными в пространстве.
Адиадохокинез характеризуется нарушением координации быстрых целенаправленных движений. При этом человек не способен выполнять быструю последовательность движений, например поочередное сгибание и разгибание пальцев. При нарушении функций мозжечка также наблюдается дезэквилибрия, т.е. потеря способности сохранять равновесие.
Мозжечок участвует не только в регуляции движений, но и осуществляет контроль над вегетативными функциями, оказывая облегчающее или угнетающее влияние на деятельность сердечно-сосудистой, дыхательной, пищеварительной систем, на терморегуляцию. Влияния мозжечка осуществляются, по-видимому, благодаря его связям с ретикулярной формацией и гипоталамусом. Деятельность мозжечка протекает в непосредственной связи с корой больших полушарий и под её контролем.
5.17. Промежуточный мозг
Лекция 3.
РЕТИКУЛЯРНАЯ ФОРМАЦИЯ
Любая ответная реакция организма, любой рефлекс представляет собой обобщенный, целостный ответ на раздражитель. В ответной реакции участвует вся ЦНС, участвуют многие системы организма. Это объединение, включение в различные рефлекторные реакции обеспечивается ретикулярной формацией (РФ). Она является главным объединителем рефлекторной деятельности всей ЦНС.
Первые сведения о РФ были получены в конце XIX и начале XX в.
Эти исследования показали, что в центральной части ствола мозга располагаются нейроны, которые имеют разную величину, форму и тесно переплетены друг с другом своими отростками. Так как внешний вид нервной ткани этой области под микроскопом напоминалсеть, то Дейтерс, который впервые описал ее строение в 1885 году, назвал ее ретикулярной или сетчатой формацией. Дейтерс считал, что РФ выполняет чисто механическую функцию. Он рассматривал ее как каркас, как арматуру ЦНС. Истинные функции РФ, физиологическое значение ее было выяснено сравнительно недавно, последние 20-30 лет, когда в руках физиологов появилась микроэлектродная техника и используя стереотаксическую методику стало возможным изучать функции отдельных участков ретикулярной формации.
Ретикулярная формация является надсегментарным аппаратом мозга,
ЦНС. Она связана со многими образованиями ЦНС.
Ретикулярная формация (РФ) образована совокупностью нейронов, расположенных в его центральных отделах как диффузно, так и в виде ядер.
Структурные особенности РФ . Нейроны РФ имеют длинные маловетвящиеся дендриты и хорошо ветвящиеся аксоны, которые часто образуют Т-образное ветвление: одна из ветвей аксона имеет нисходящее, а вторая - восходящее направления. Ветви нейронов под микроскопом образуют сеточку (ретикулум), с чем и связано название данной структуры мозга, предложенное О. Дейтерсом (1865).
Классификация.
1 . С анатомической точки зрения РФ делят на:
1. Ретикулярную формацию спинного мозга - это substantioРоланди, которая занимает верхушку задних рогов верхних шейных сегментов.
2. Ретикулярную формациюствола мозга (заднего и среднего мозга).
3. Ретикулярную формацию промежуточного мозга. Здесь она представлена неспецифическими ядрами таламуса и гипоталамуса.
4. Ретикулярную формацию переднего мозга.
2. В настоящее время физиологи пользуются классификацией РФ, которую предложил шведский нейрофизиолог Бродал. Согласно этой классификации в РФ выделяют латеральное и медиальное поля .
Латеральное поле - это афферентная часть РФ. Нейроны латерального поля воспринимают информацию, которая приходит сюда, поступает по восходящим и нисходящим проводниковым путям. Дендриты этих нейронов направлены латералъно и они воспринимают сигнализацию. Аксоны идут в сторону медиального поля, т.е. обращены в центр мозга.
Афферентные входы поступают в латеральные области РФ преимущественно от трех источников:
Температурных и болевых рецепторов по волокнам спиноретикулярного тракта и тройничного нерва. Импульсы идут в ретикулярные ядра продолговатого мозга и моста;
Сенсорной, от зон коры головного мозга по кортико-ретикулярным путям идут в ядра, дающие начало ретикулоспинальным трактам (гигантоклеточное ядро, оральное и каудальное ядра моста), а также в ядра, которые проецируются на мозжечок (парамедианное ядро и ядро покрышки моста);
Ядер мозжечка по мозжечково-ретикулярному пути импульсация поступает в гигантоклеточное и парамедианное ядра и ядра моста.
Медиальное поле - это эфферентная, исполнительная часть РФ. Оно расположено в центре мозга. Дендриты нейронов медиального поля направлены в сторону латерального поля, где они контактируют с аксонами латерального поля. Аксоны нейронов медиального поля идут либо вверх, либо вниз, образуя восходящие и нисходящие ретикулярные пути. Ретикулярные пути, которые образованы аксонами медиального поля образуют широкие связи со всеми отделами ЦНС, спаивая их воедино. В медиальном поле формируются преимущественно эфферентные выходы.
Эфферентные выходы идут:
К спинному мозгу по латеральному ретикулоспинальному тракту (от гигантоклеточного ядра) и по медиальному ретикулоспинальному тракту (от каудального и орального ядер моста);
К верхним отделам головного мозга (неспецифическим ядрам таламуса, заднему гипоталамусу, полосатому телу) идут восходящие пути, начинающиеся в ядрах продолговатого мозга (гигантоклеточном, латеральном и вентральном) и в ядрах моста;
К мозжечку идут пути, которые начинаются в латеральном и парамедианном ретикулярных ядрах и в ядре покрышки моста.
Медиальное поле в свою очередь делят на восходящую ретикулярную систему (ВРС) и нисходящую ретикулярную систему (НРС) . Восходящая ретикулярная система образует проводящие пути, направляет свои импульсы коре больших полушарий и подкорке. Нисходящая ретикулярная система направляет свои аксоны в нисходящем направлении - в спинной мозг - ретикуло-спинальный путь.
Как в восходящей, так и нисходящей ретикулярной системе имеются тормозные и активирующие нейроны. По этому различают восходящую ретикулярную активирующую систему (ВРАС) , и восходящую ретикулярную тормозную систему (ВРТС) . ВРАС оказывает активирующее влияние на кору и подкорку, а ВРТС - тормозит, подавляет возбуждение. Так же в НРС различают нисходящую ретикулярную тормозную систему (НРТС) которая берет начало от тормозных нейронов РФ и идет в спинной мозг тормозит его возбуждение, и нисходящую ретикулярную активирующую систему (НРАС), которая направляет активирующие сигналы в нисходящем направлении.
Функции ретикулярной формации
Ретикулярная формация не осуществляет специфические, какие-то определенные рефлексы Функция РФ иная.
1. Во-первых, РФ обеспечивает, интеграцию, объединение функций всей ЦНС. Она является главной интегрирующей, ассоциативной системой ЦНС. Эту функцию она выполняет потому, что РФ, ее нейроны образуют огромное количество синапсов как между собой, так и с другими отделами ЦНС. Поэтому, возбуждение попав в РФ, очень широко распространяется, иррадиирует и по ее эфферентным путям: восходящим и нисходящим это возбуждение достигает всех частей ЦНС. В результате этой иррадиации включаются и вовлекаются в работу все образования ЦНС, достигается содружественная работа отделов ЦНС т.е. РФ обеспечивает формирование целостных рефлекторных реакций , в рефлекторной реакции участвует вся ЦНС
II. Вторая функция РФ заключается в том, что она поддерживает тонус ЦНС, т.к. РФ сама всегда находится в тонусе, тонизирована. Её тонус обусловлен рядом причин.
1).РФ обладает очень высокой хемотропностью Здесь находятся нейроны обладающие высокой чувствительностью к некоторым веществам крови (например, к адреналину, СО;) и лекарствам (к барбитуратам, аминазину и др.).
2). Второй причиной тонуса РФ является то, что в РФ постоянно поступают импульсы от всех проводниковых путей. Это связано с тем, что на уровне ствола мозга афферентное возбуждение, которое возникает при раздражении любых рецепторов трансформируется вдва потока возбуждения. Один поток направляется по классическому лемнисковому пути, по специфическому пути и достигает определенную для данного раздражения участок коры. Одновременно каждый проводниковый путь по коллатералям отклоняется в РФ и возбуждает её. Не все проводниковые пути действуют одинаково на тонус РФ. Возбуждающее действие проводниковых путей неодинаково. Особенно сильно возбуждает РФ импульсы, сигналы, которые идут от болевых рецепторов, с проприорецепторов, со слуховых и зрительных рецепторов. Особенно сильно возбуждения возникает при раздражении окончаний тройничного нерва. Поэтому при обмороках раздражают окончания n. trigeminus : обливают водой, дают нюхать нашатырный спирт, (йоги, зная действие тройничного нерва, устраивают "прочищение мозгов" - делают несколько глотков воды через нос).
3). Тонус РФ поддерживается также за счет импульсов, которые идут по нисходящим путям от коры больших полушарий, от базальных ганглиев.
4). В поддержании тонуса сетчатого образования имеет большое значение такжедлительная циркуляция нервных импульсов в самой РФ, имеет значение реверберация импульсов в РФ. Дело в том, что в РФ находится громадное количество нейронных колец и по ним информация, импульсы циркулируют часами.
5). У нейронов РФ длительный латентный период ответа на периферическую стимуляцию в связи с проведением возбуждения к ним через многочисленные синапсы.
6). Они имеют тоническую активность, в покое 5-10 имп/с.
В результате выше названных причин РФ всегда находится в тонусе и от нее импульсы поступают в другие отделы ЦНС. Если перерезать ретикуло-кортикальные пути, т.е. восходящие пути, идущие от РФ в кору, то кора головного мозга выходит из строя, так как она лишилась главного источника импульсов.
Похожая информация.
Ретикулярная формация
(от лат. reticulum - сеточка, formatio - образование)
сетевидное образование, совокупность нервных структур, расположенных в центральных отделах стволовой части мозга (продолговатом и среднем мозге, зрительных буграх). Нейрон ы,
составляющие Р. ф., разнообразны по величине, строению и длине Аксон ов;
их волокна густо переплетаются. Термин «Р. ф.», введённый немецким учёным О. Дейтерсом, отражает лишь морфологические её особенности. Р. ф. морфологически и функционально связана со спинным мозгом, мозжечком (См. Мозжечок), лимбической системой (См. Лимбическая система) и корой больших полушарий головного мозга. В области Р. ф. осуществляется взаимодействие поступающих в неё как восходящих - афферентных, так и нисходящих - эфферентных импульсов. Возможна также циркуляция импульсов по замкнутым нейронным цепям. Т. о., существует постоянный уровень возбуждения нейронов Р. ф., вследствие чего обеспечиваются тонус и определённая степень готовности к деятельности различных отделов центральной нервной системы. Степень возбуждения Р. ф. регулируется корой больших полушарий головного мозга (См. Кора больших полушарий головного мозга).
Нисходящие влияния. В Р. ф. различают области, которые оказывают тормозящие и облегчающие влияния на двигательные реакции спинного мозга (См. Спинной мозг) (рис. 1
). Зависимость между раздражением различных областей ствола мозга и спинномозговыми рефлексами впервые отметил в 1862 И. М. Сеченов .
В 1944-46 американский нейрофизиолог Х. Мэгоун с сотрудниками показали, что раздражение различных участков Р. ф. продолговатого мозга оказывает облегчающее или тормозящее влияние на двигательные реакции спинного мозга. Электрическое раздражение медиальной части Р. ф. продолговатого мозга у наркотизированных и децеребрированных кошек и обезьян сопровождается полным прекращением движений, вызываемых как рефлекторно, так и стимуляцией двигательных участков коры мозга. Все тормозные эффекты - двусторонние, но на стороне раздражения такой эффект нередко наблюдается при более низком пороге раздражения. Некоторые проявления тормозящих влияний Р. ф. продолговатого мозга соответствуют картине центрального торможения, описанного Сеченовым (см. Сеченовское торможение). Раздражение латеральной области Р. ф. продолговатого мозга по периферии области, оказывающей тормозящие влияния, сопровождается облегчающим действием на моторную активность спинного мозга. Область Р. ф., оказывающая облегчающие влияния на спинной мозг, не ограничивается продолговатым мозгом, а распространяется кпереди, захватывая область варолиева моста и среднего мозга. Р. ф. может воздействовать на различные образования спинного мозга, например на альфа-мотонейроны, иннервирующие основные (экстрафузальные) волокна мышц, участвующих в произвольных движениях. Увеличение латентных периодов ответов мотонейронов при раздражении тормозящих отделов Р. ф. позволяет предполагать, что тормозящие влияния ретикулярных структур на двигательные реакции спинного мозга осуществляются с помощью вставочных нейронов, возможно клеток Реншоу. Механизм влияния Р. ф. на мышечный тонус раскрыт шведским нейрофизиологом Р. Гранитом, показавшим, что Р. ф. влияет также на активность гамма-мотонейронов, аксоны которых идут к так называемым интрафузальным мышечным волокнам, осуществляя важную роль в регуляции позы и фазных движений организма. Восходящие влияния. Различные отделы Р. ф. (от промежуточного до продолговатого мозга) оказывают возбуждающие генерализованные влияния на кору мозга, т. е. вовлекают в процесс возбуждения все области коры больших полушарий (рис. 2
). В 1949 итальянский физиолог Дж. Моруцци и Мэгоун, исследуя биоэлектрическую активность мозга, установили, что раздражение Р. ф. ствола мозга изменяет медленные синхронные высоковольтные колебания, характерные для сна, на низкоамплитудную высокочастотную активность, характерную для бодрствования. Изменение электрической активности коры мозга сопровождается у животных внешними проявлениями пробуждения. Р. ф. тесно связана анатомически с классическими проводящими путями, и возбуждение её осуществляется с помощью экстеро- и интероцептивных афферентных (чувствительных) систем. На этом основании ряд авторов относит Р. ф. к неспецифической афферентной системе мозга. Однако применение различных фармакологических веществ при изучении функции Р. ф., открытие избирательного действия химических препаратов на реакции, осуществляемые с участием Р. ф., позволили П. К. Анохин у
сформулировать положение о специфичности восходящих влияний Р. ф. на кору мозга. Активирующие влияния Р. ф. всегда имеют определённое биологическое значение и характеризуются избирательной чувствительностью к различным фармакологическим веществам (Анохин, 1959, 1968). Введённые в организм наркотические средства вызывают торможение нейронов Р. ф., блокируя тем самым её восходящие активирующие влияния на кору мозга. Важная роль в поддержании активности Р. ф., чувствительных к различным циркулирующим в крови химическим веществам, принадлежит гуморальным факторам: катехоламинам, двуокиси углерода, холинэргическим веществам и т. д. Это обеспечивает включение Р. ф. в регулирование некоторых вегетативных функций. Кора больших полушарий, испытывающая тонические активирующие влияния со стороны Р. ф., может активно изменять функциональное состояние ретикулярных образований (изменять скорость проведения возбуждения в ней, влиять на функционирование отдельных нейронов), т. е. контролировать, по выражению И. П. Павлова, «слепую силу» подкорки. Открытие свойств Р. ф., её взаимоотношений с другими подкорковыми структурами и областями коры мозга позволило уточнить нейрофизиологические механизмы боли, сна, бодрствования, активного внимания, формирования целостных условно-рефлекторных реакций, развития различных мотивационных и эмоциональных состояний организма. Исследования Р. ф. с использованием фармакологических средств открывают возможности медикаментозного лечения ряда заболеваний центральной нервной системы, обусловливают новый подход к таким важнейшим проблемам медицины, как наркоз и др. Лит.:
Бродал А., Ретикулярная формация мозгового ствола, пер, с англ., М., 1960; Росси Дж. Ф., Цанкетти А., Ретикулярная формация ствола мозга, пер. с англ., М., 1960; Ретикулярная формация мозга, пер. с англ., М., 1962; Мэгун Г., Бодрствующий мозг, пер. с англ., 2 изд., М., 1965; Анохин П. К., Биология и нейрофизиология условного рефлекса, М., 1968; Гранит Р., Основы регуляции движений, пер. с англ., М., 1973; Moruzzi G., Magoun Н. W., Brain stem reticular formation and activation of EEG, в книга: Electroencephalography and clinical neurophysiology, v. 1, Boston, 1949 В. Г. Зилов.

Большая советская энциклопедия. - М.: Советская энциклопедия . 1969-1978 .
Смотреть что такое "Ретикулярная формация" в других словарях:
- (formatio reticularis; лат. reticulum сеть; синоним ретикулярная субстанция) комплекс клеточных и ядерных образований, занимающих центральное положение в стволе головного мозга и в верхнем отделе спинного мозга. Большое количество нервных волокон … Википедия
Ретикулярная формация - Сложная сеть нейронов и клеточных ядер, занимающая центральную часть ствола головного мозга. Часто называется «ретикулярной системой активации» из за той роли, которую она играет в процессе активации. Совзременные исследования позволяют… … Большая психологическая энциклопедия
Совокупность структур в центральных отделах головного мозга, регулирующих уровень возбудимости и тонуса ниже и вышележащих отделов центральной нервной системы, включая кору больших полушарий … Большой Энциклопедический словарь
РЕТИКУЛЯРНАЯ ФОРМАЦИЯ, сложный механизм ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ позвоночных, находящийся в стволе спинного МОЗГА. Состоит из взаимосвязанных скоплений тел нервных клеток (серое вещество) и считается, что влияет на многие физиологические… … Научно-технический энциклопедический словарь
- (formatio reticularis), совокупность нервных структур, расположенных в спинном, продолговатом, среднем мозге и варолиевом мосту и образующих единый функц. комплекс. Филогенетически древняя система двигат. контроля. Хорошо развита у всех… … Биологический энциклопедический словарь
Ретикулярная формация - (лат. rete сеть, formatio формирование, образование, составление) сетевидная нервная структура, состоящая из более чем 50 ядер и обширной сети нейронов со сложными и разветвлёнными аксональными и дендритными отростками. Название предложено… … Энциклопедический словарь по психологии и педагогике
Совокупность структур, расположенных в спинном, продолговатом и среднем мозге и варолиевом мосту и образующих единый функциональный комплекс. Оказывает активирующее и тормозящее влияние на различные отделы центральной нервной системы, повышая… … Энциклопедический словарь
ретикулярная формация - (formatio reticularis) совокупность небольших, но многочисленных ядер, расположенных в центральных отделах ствола мозга. Нейроны ретикулярной формации имеют сильно ветвящиеся отростки, идущие в различных направлениях, напоминая под микроскопом … Словарь терминов и понятий по анатомии человека
Ретикулярная формация - (от лат. reticulum сетка) нервная структура, расположенная вдоль всего мозгового ствола и состоящая из клеток, отростки которых ветвятся в обширных областях мозговой коры. Функция ретикулярной формации заключается в активизации коры головного… … Психология человека: словарь терминов
I Ретикулярная формация (formatio reticularis; лат. reticulum сетка; синоним ретикулярная субстанция) комплекс клеточных и ядерных образований, занимающих центральное положение в стволе головного мозга и в верхнем отделе спинного мозга. Большое… … Медицинская энциклопедия